Zašto su mtDNA i Y-kromosomske loze važne?

(Kako će ići slijedećih nekoliko tekstova o nekim čudnim genetskim podudarnostima, ali i "zaboravljenim" starim narodima svijeta, koji zapravo još uvijek postoje - moram objasniti te halpogrupe. Zato ovaj malo podulji tekst o, zapravo, genetici. I kako se to sve gleda kao isključivi dokaz porijekla naroda u današnja vremena)
U današnje vrijeme, u svakoj regionalnoj ili populacijskoj povijesti nailazima na reference prema Y-kromosomskoj i mtDNK "haplogrupi". Iako nikada nisu u središtu priče (iako nekako sada i jesu), to nisu ukrasi, niti su trivijalnosti. Haplogrupe dolaze uz nejasnu nomenklaturu, kombinacijom slova i brojeva, nepredvidljive su duljine. Ali, jednostavnije se njihovi međusobni odnosi mogu vizualizirati kao doslovna shema, nalik razgranatom stablu. Također, uklapaju se u narativne strukture, nekada. Npr. reći kako su "R1b i R1a - 'bratske' haplogrupe" jest nešto što razumije velika većina ljudi, umjesto kada se pokuša objasniti: "'populacija duh' koja je dodana modelu - rješava f-statistiku u ukupnom grafu populacije"?! Haplogrupe jesu dosta korisni putokazi, ali u eri sveprisutnog GPS-a, možda su malo više poput dragocjenih fragmenata iz ranih karata povijesnih društava, nego što se tiče preciznih satelitskih koordinata.
Studije Y i mtDNK su genomski miljenici prijelaza ovog mladog stoljeća, ali sada je ocvjetalo, dok jurimo dublje u postgenomsku eru. Ima li uistinu smisla promatrati ove krhke i male vitice genealoških arkana, sada kada se sve tri milijarde ljudskih baznih parova mogu rutinski osvijetliti? Paradoksalno, ali upravo ta činjenica što više nisu glavne zvijezde showa - oslobodilo je Y i mtDNA markere da ostvare svoj puni i jedinstveni potencijal, u složenim narativima povijesne populacijske genetike. Nikada nisu imali dosegli pun smisao kao idoli matineja. Više su bile poput karakternih glumaca koji ukradu pojedine scene, i sada su konačno došli do izražaja; sada kada su se iskoristili neka osvijetle različite demografske povijesti spolova zbog kulture (poliginija ili poliandrija). ili zbog osvajanja (hvatanje žena starosjedilaca), ili de precizno poprate loze visokog statusa kroz vrijeme. Danas su ovi iznimni alati konačno dobili vrijednu misiju. Nakon što analize cijelog genoma uokvire širi narativ, Y i mtDNA loze mogu razraditi specifične detalje, poput spolne demografije ili slabih veza između populacija, a koje se ne mogu uočiti drugim metodama.
Y i mtDNK loze su tehnički uniparentalni markeri, jer se prenose samo sa jednog roditelja - Y sa oca na sina, a mtDNK sa majke na dijete (iako sinovi ne prenose svoju mtDNK na svoje potomstvo, i tu su to slijepe ulice). Dakle, informacije koje prenose su vrlo rodbinski bliske. Unutar kruga genetske genealogije: izravne očinske i majčine loze osobe imaju neporecivu emocionalnu vrijednost, koja im donosi određeni neupitni primat - iako te uske linije podrijetla nisu jedinstveno privilegirane, u bilo kojem znanstvenom smislu. Y i mtDNK loze isključivo prate vrlo specifične i precizne pretke, unutar određenog obiteljskog stabla. Ako upoznam Poljaka koji nosi haplogrupu R1a1a, onda mogu sa sigurnošću zaključiti kako dijelimo nekog izravnog zajedničkog pretka, od otprilike prije 5500 godina, i koji je živio negdje na pontskoj stepi Ukrajine. Oko 220 generacija očeva i sinova se proteže sve do jednog čovjeka, izravnog očevog pretka, pretka otprilike trećine slavenskih muškaraca.
Haplogrupe su način "rezbarenja prirode u zglobu", duž poznatih linija, koje se ljudima čine prirodnima i pristupačnima. Svaka pojedina Y haplogrupa prati izravno očevo podrijetlo, duž dugog lanca muškaraca, unatrag do podrijetla naše vrste (podaci su postojali i ranije); dok je mtDNK haplogrupa genealogija svih naših majki, unatrag do evokativne "mitohondrijske Eve". Ove jednostavne i izravne genetske linije se, prikladno, temelje na istim pretpostavkama kao i naše kulturne tradicije iz prezimena, te vjerno odražavaju fiksaciju naše vrste na patrilinearno i matrilinearno podrijetlo. Obje linije prate taj mali trag nečijeg podrijetla, isključivo duž izravnih genealogija jednog spola, baš kao što to čine Y kromosom i mtDNK. Unutar konteksta ljudske demografske povijesti, očinska i majčina dinamika variraju na zanimljive načine, koje mogu otkriti distribucije haplogrupa i filogenetska stabla u koja su ugrađene. Nevjerojatno je, iako nije iznenađujuće, npr. potvrditi kako su, u rasno miješanim populacijama Kolumbije, gotovo sve mtDNK haplogrupe doista autohtone za Ameriku, dok Y kromosomi izgledaju kao poduzorak iberijskih muškaraca.
Možemo jednostavno reći: mtDNK i Y studije dodaju kvantitativnu dimenziju kvalitativnom znanju.
Danas Y i mtDNK loze igraju sporednu ulogu pri analizama cijelog genoma, u konstelaciji uvida koje možemo prikupiti o našoj evolucijskoj i genetskoj prošlosti. Budući da danas znamo puno više, pomognuti alatima 21. stoljeća, poput sekvenciranja cijelog genoma i paleogenetike polja - ova dva markera sada mogu istinski doći do izražaja - kako bi istražili vrlo precizne demografske pojave, umjesto da se nerealno koriste za osvjetljavanje cjelokupnosti ljudske demografske prošlosti.
Haplotip
Dakle, što je haplogrupa? To je zapravo samo skup srodnih haplotipova, koji sežu do zajedničkog pretka unutar genealogije. U redu, a što je onda haplotip?
Slijed ljudske DNK dolazi u dvije kopije, jedan genom od tri milijarde baza koji je dupliciran u dva lanca, zapakiran i organiziran u kromosome. To znači: svaki naš gen ima dvostruku redundanciju - uz ključne iznimke spolnih kromosoma kod muškaraca i mitohondrijske DNK kod svih nas. Svaka pojedina pozicija u genomu, kada se zatraži unos, okreće se i savjetuje sa partnerskom pozicijom, prije nego registrira svoj glas ili donese odluku. Osim svake ovakve usamljene i male pozicije (kojih ima više od 50 milijuna) na malom Y kromosomu svakog muškarca, postoje i pozicije na sićušnim, ali moćnim motorima, koji pokreću svaku našu stanicu, tj. našim mitohondrijima (mnogo kraći od Y, sa samo 16569 pozicija), pri čemu su ovi potonji naslijeđeni od izravne linije ženskih predaka svakog djeteta, i to pri njihovom začeću.
Osim što u genomu postoje milijarde pozicija koje se poravnavaju u nizu, geni unutar organizama su raspoređeni određenim redoslijedom i združeni su u specifične kromosome. Oni ne plutaju slobodno u jezgri, nego su upakirani unutar precizne strukture. Što je veća evolucijska udaljenost koja razdvaja dvije vrste, veća je vjerojatnost kako će se razlikovati u redoslijedu svojih gena (i ukupnom broju svojih kromosoma; ljudi imaju 46 kromosoma, dok naši navodni rođaci, veliki majmuni ih imaju 48). Nasuprot tome, unutar vrsta općenito postoji isti redoslijed gena.
Ali, i u tim uređenim genima i dalje postoji bezbroj varijacija. No, upravo varijabilne pozicije, jednonukleotidni polimorfizmi (SNP), mnoštvo A, C, G i T gena, kada su grupirani zajedno u određenom slijedu i korelirani u svom nasljeđivanju kao blok - karakteriziraju haplotip. Haplotip je definiran povezanom genetskom varijacijom na segmentu DNA, naslijeđenom kao jedinica od roditelja. Donja slika ilustrira dva kratka slijeda koja su gotovo identična, ali se razlikuju na tri pozicije: 4, 16 i 27. Haplotip 1 se čita kao C, T i C na tim pozicijama; dok se haplotip 2 čita kao G, G i A. Budući da svi geni, osim onih na muškom spolnom kromosomu, dolaze u dvije kopije, možete vidjeti kako pojedinac zapravo može nositi i haplotip 1 i haplotip 2:

U većem dijelu ljudskog genoma, ovi nizovi povezanih SNP varijacija postaju fragmentirani i odvojeni jedni od drugih, tijekom generacija zbog genetske rekombinacije, jer se segmenti DNK lome i križaju, kako bi se podudarali sa alternativnom kopijom unutar vašeg genoma. Zamislite, npr. jedna osoba kineskog podrijetla se uda za osobu iz malog švedskog sela; u toj prvoj generaciji njihovih potomaka, genetski različiti preci kineskog roditelja bili bi grupirani duž dugih dijelova DNK (parovi kromosoma bili bi isključivo kineski ili isključivo švedski, jedan homolog od svakog roditelja). Ali, u svakoj sljedećoj generaciji, u kojoj njihovi potomci ostanu u Švedskoj miješajući se i dalje sa Šveđanima, segmenti predaka kineskog podrijetla će biti razdvojeni i rekombinirani sa ovom DNK europskog podrijetla. Nakon mnogo stoljeća, tragovi izvornih dugih kineskih haplotipova povezanih varijacija će biti moguće detektirani samo kao puno kraći segmenti.
Ovaj mehanizam možemo ilustrirati u jednoj generaciji, uz gore navedene haplotipove, ako ovi zamijene nizove nakon prekida; u ovom slučaju između jedanaestog i dvanaestog slova:

Haplotipovi su sada CGA i GTC, umjesto CTC i GGA.
Kada muškarci i žene formiraju spermije i jajne stanice tijekom mejoze, očekujemo oko 20-40 događaja 'crossover'-a, zamjenjujući segmente preko parova kromosoma, naslijeđenih od majke i oca pojedinca (kao i X kromosoma kod žena). Ovi prekidi i zamjene između parova kromosoma se mogu nakupljati tijekom generacija, smanjujući povezanost genetskih varijanti samo preko segmenata DNK, usitnjavajući vrlo duge haplotipove. Ova trajna dinamika, a koja djeluje protiv povezanih varijacija, razlog je zašto prisutnost vrlo dugih, ali i uobičajenih haplotipova, često zaslužuje detaljnije ispitivanje. Npr. među većinom Europljana i velikom manjinom Južnih Azijata, jedan od najdužih haplotipova je pronađen u genu laktaze, segmentu koji je dugačak preko milijun baza (to bi bilo 1,7% duljine Y kromosoma).
Zašto je ovaj haplotip toliko i dug i toliko čest? Svoju ogromnu popularnost duguje činjenici koja uključuje SNP mutaciju, unutar laktaze koja daje toleranciju na laktozu, osobinu koja je bila pod vrlo jakom prirodnom selekcijom i među stočarima koji piju mlijeko, tijekom posljednjih 3000 godina. Ako se frekvencija preferiranog SNP-a dovoljno brzo povećava zbog jake pozitivne selekcije, tada će se slučajno i susjedni dijelovi genoma "autostopirati", nadjačavajući uobičajeni spori, ali neumoljivi napredak rekombinacije, ostavljajući vrlo dugi haplotip, sa vrlo visokom frekvencijom u populaciji.
Otkrivanje dugog haplotipa može biti mogući znak kako je primarna evolucijska sila, tj. prirodna selekcija, gazila kroz ljudski genom, ostavljajući za sobom ogromnu pustinju genetskih varijacija. Haplotip oko laktaze je kopija samo jedne jedinke, koja je vjerojatno živjela prije oko 5000 godina u središnjoj Euroaziji, tako da u većini sjevernoeuropskih genoma uopće niti nema nikakve razlike između njihove dvije naslijeđene kopije kromosoma.

Ovo su polovice prvih devet kromosoma koje je kćer naslijedila od oca. Magenta segmenti su od očevog oca, dok su sivi od očeve majke. Točke na kojima kromosomi prelaze iz jedne boje u drugu ukazuju na događaje rekombinacije, koji su se dogodili tijekom mejoze. Možete vidjeti kako samo kromosom 6 ne pokazuje događaj rekombinacije.
Ali, vratimo se na mjesto gdje je rečeno kako se u većini genoma haplotipovi raspadaju. Naša današnja priča niti nije o ogromnom dijelu genoma, gdje se odvija sve to ogromno prenošenje, rekombiniranje i križanje. Za haplotipove, naši informatori su sićušni dijelovi genoma, koji koračaju manje utabanim putem. I takva njihova samostalna putovanja su napravila svu potrebnu razliku. Mitohondrijski genom i većina Y kromosoma su (prema namjeri) samostalni plesači, koji se pojavljuju netaknuti (u svakom činu) sve dok evolucija napreduje, nikada ne surađuju, niti se dogovaraju ni sa kim (postoje uvijek neke iznimke, ali one nisu relevantne za ovu raspravu). Na Y i mtDNK, mutacije koje nastaju zajedno i ostaju zajedno, baš poput mnogih grafita koji se vremenom skupe na zidu, jer nikada nema rekombinacije, koja bi ih zamijenila za drugi segment. Jedini način na koji se sekvence mtDNK i Y mijenjaju vremenom jest kada akumuliraju nove mutacije. Nema miješanja, niti podudaranja varijanti. Dok će, npr. naši kromosomi 1 vjerojatno biti mješavina naših četvero baka i djedova zbog rekombinacije kod roditelja - mtDNK je jednostavno ona vaše majke, i njene majke, i njene majke prije nje. Muški Y kromosom je jednostavno ona njegovog oca, njegovog oca prije njega, itditd.
Evolucijsku povijest haplotipova, u većem dijelu genoma, bilo bi teško rekonstruirati zbog komplicirane varijable rekombinacije koja miješa segmente, stvarajući mnoštvo varijacija haplotipova, tijekom mnogo generacija. Srećom, za naše potrebe, ova diskvalificirajuća komplikacija se ne odnosi na Y kromosom i mtDNK. Umjesto toga, oni su sami po sebi pojedinačni haplotipovi, od početka do kraja, koji akumuliraju varijacije kroz mutacije, tijekom generacija. Jednostavna evolucijsko-genetska dinamika Y i mtDNK sama po sebi olakšava demografsko zaključivanje, jer samo trebate usmjeriti obiteljsko stablo do zajedničkih predaka.

Haplogrupa
Dakle, ako je haplogrupa samo skup srodnih haplotipova koji dijele zajedničkog pretka, znači li to da smo svi dijelom opće ljudske haplogrupe? Da, jer u kontekstu dva tipa uniparentalnih loza, Y i mtDNA filogenija, one se naravno spajaju sve do najnovijih zajedničkih predaka, utjelovljenih u mitskim pojedincima koje nazivamo mitohondrijskom Evom i Y-kromosomskim Adamom. Unatoč tim evokativnim imenima, rad sa uniparentalnim lozama znači biti zauvijek osuđen na naglašavanje kako ove dvije osobe nisu bile jedini ljudi koji su živjeli u svojoj generaciji, i nisu nužno doprinijele nesrazmjernom količinom svog podrijetla ljudima koji danas žive. Samo što će, prema matematici uniparentalnog nasljeđivanja, na kraju sve konkurentske neprekinute loze (osim jedne) izumrijeti, tako da će jedna linija, koja potječe od zajedničkog pojedinačnog pretka obuhvatiti cijelu populaciju. Možda je slučajno, ali je i neizbježno.
Logika ovoga nije jedinstvena samo za DNK. Odražava se u našim kulturnim i genealoškim tradicijama. Prije gotovo 2000 godina, car August je bio zabrinut zbog propadanja i izumiranja rimskih plemićkih obitelji. One su bile organizirane u gensove, potomke zajedničkih muških predaka i čiji se status nasljeđivao lancem legitimnih patrilinearnih potomaka. Ono što August nije razumio jest da problem nije bio moral plemstva, već običaj patrilinearnosti u službeno monogamnoj kulturi, budući mnogi muškarci u svakoj generaciji nisu imali legitimne žive sinove, kako bi nastavili svoju obiteljsku lozu. August je žudio za pouzdanom plodnošću, kao i bezbrižnim nedostatkom brige o očinstvu, kao kod zečeva, u svijetu gdje je težnja za postizanjem očinstva više nalikovala ožujskom ludilu, koje se urušavalo generaciju po generaciju. Kao tragičan primjer, car Marko Aurelije imao je najmanje sedam legitimnih sinova, ali samo je jedan, ludi Komod, preživio do odrasle dobi (i na kraju su njegove ludorije dovele do toga da ga uviju u atentatu). U patrilinearnom ili matrilinearnom sustavu neophodno je potrebna neprekinuta linija muškaraca ili žena, ali priroda vjerojatnosti znači kako će neke loze izumrijeti u svakoj generaciji, čime se taj slijed muškaraca ili žena konačno prekida.

Lucije Kornelije Sula Feliks, reakcionarni rimski diktator, slavno je našalio: "Nijedan prijatelj mi nikada nije služio, niti mi je neprijatelj ikada učinio nepravdu, a da mu nisam u potpunosti odužio."
Ali, što je s onim obiteljima koje su procvjetale i postale još plodnije? U Rimu su se različite grane, posebno ako su bile istog i brojnog roda, funkcionalno razdvajale. Tijekom kasne Republike, diktator Lucije Kornelije Sula dao je ubiti senatora Lucija Kornelija Cinu, jer je ovaj podržavao neprijatelje prvog. Iako su obojica bili iz roda Kornelija, grane Cina i Sula, do prvog stoljeća prije Krista, smatrane su potpuno odvojenim obiteljima, bez posebno jake veze. U jeziku haplogrupa: Korneli su bili makro-haplogrupa, dok su Cina i Sula bili potomci tih haplogrupa. Kako neke haplogrupe izumiru, druge se diverzificiraju kako bi popunile prazninu, tako da će se na kraju ispostaviti kako sve haplogrupe potječu od zajedničkog pretka, baš kao i Korneli:
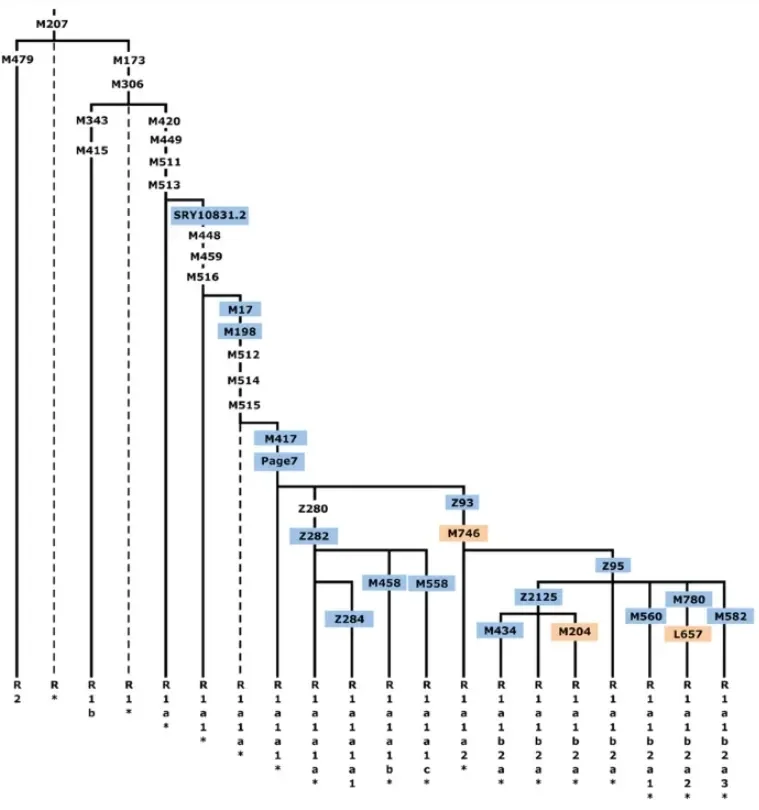
Budući da Y i mtDNA sekvence gomilaju mutacije kako napreduju kroz vrijeme, njihove filogeneze definirane su novim mutacijama, koje se akumuliraju unutar njihovih obiteljskih stabala, označavajući značajnu točku grananja. U nekom određenom trenutku, u dalekoj prošlosti, sve loze dijelile su iste zajedničke mutacije u posljednjem zajedničkom pretku, ali tijekom generacija, dolazile su nove varijante koje su definirale ovu ili onu granu. Ako je jedna određena loza bila reproduktivno uspješna, izrodila bi mnoge manje grane, koje su od nje potekle.
Gore je slika koja definira grane Y-kromosomske haplogrupe R koje se razlikuju po brojnim mutacijama, a koje su se akumulirale tijekom desetaka tisuća godina (najbolja procjena jest kako su se R1 i R2 razdvojili prije oko 35000 do 40000 godina). Iako postoje različite nomenklature za Y i mtDNA haplogrupe, ovaj zajednički sustav je prilično jednostavan, u načinu na koji se imena prate uz mutacijame. Prvo, dvije mutacije, M479 i M173, definiraju razdvajanje između R1 i R2. Zatim, unutar R1 vidite mutacije koje definiraju R1b i R1a; i slijedeće unutar R1a vidite mutaciju M17, koja definira R1a1a. Pproces ide dalje unutar R1a u ovom stablu, a neki genealoški fanatici definiraju jedinstvene mutacije, koje prate bliske grane iste obitelji.
Postoje slučajevi, kao u mtDNA haplogrupi U (slika u nastavku), gdje su se mutacije dogodile toliko blizu u vremenu, te ne možemo definirati bifurkacije, pa toliko grana izlazi iz jednog čvora:

Konačno: samo zato što mutacije definiraju određenu granu, to ne znači kako sve one imaju istu važnost za nas. Unutar R1a1a, mutacije Z282 i Z93 zadržavaju izvanrednu važnost, jer definiraju loze koje obuhvaćaju veliku većinu europskih i azijskih muškaraca ove haplogrupe, i povezane su sa masovnim demografskim širenjem Indoeuropljana iz pontske stepe.
Što nam mtDNA i Y-kromosomske loze još uvijek mogu reći?

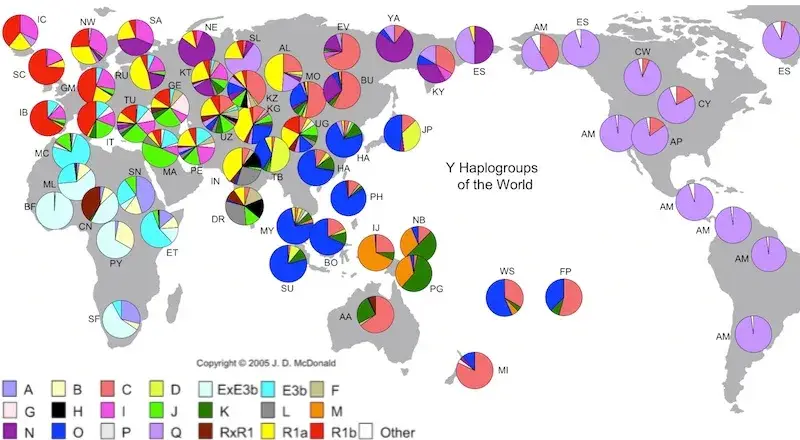
Postojeće haplogrupe Y kromosoma
Iako znanost teži otkrivanju vječnih istina, njena praksa ostaje povijesno ograničena. Prije nekoliko desetljeća, neobična sinergija bioloških stvarnosti, tehnologija i metoda se ujedinila kako bi istaknula uniparentalne loze. Prvo, mtDNK je nevjerojatno obimna, jer svaka ljudska stanica ima 1000 do 6000 mitohondrija; za razliku od jednog nuklearnog genoma. Dakle, prije nego što su tehnike lančane reakcije polimeraze omogućile amplifikaciju (čak i malih količina DNK tijekom 1990-ih), mtDNK je bila ciljana, jer ju je bilo lako izdvojiti i dobiti. Izvorni rad na mtDNK i koji je doveo do mitohondrijske Eve se oslanjao na prikupljanje odbačene posteljice bogate tkivom, koja je obilan i dostupan izvor genetskog materijala. Ali, čak i nakon pojave PCR-a, tehnologije sekvenciranja su bile izuzetno mukotrpne i skupe. Nadalje, poznato je da mtDNK ima u sebi hipervarijabilnu regiju, koja je bogata genetskim informacijama, ali i brže akumulira mutacije (koji su uvijek informativni markeri) na evolucijskoj vremenskoj skali. Dakle, mitohondrijska DNK nije izašla iz mode, čak ni kada su joj se druge genetske tehnike pridružile u demografskom zaključivanju.
Iako je Y kromosom imao manje urođenih prednosti od mtDNK (bilo je puno teže izdvojiti dovoljne količine, a uzorci nisu imali toliku gustoću varijacija kao mtDNK), bio je siromašan funkcionalnim genima. Stoga se pretpostavljalo kako ga nije snažno oblikovala prirodna selekcija, što ga je učinilo savršenim tragačem za demografskom povijesti. Y-kromosom, baš poput mtDNK, bilježi kako se odnosite prema drugima, a ne kako se prilagođavate svojoj okolini. Konačno, nijedan marker se ne rekombinira, što znači da su vrlo lako obradivi za modeliranje na računalnim platformama, koji su postojeli prije jedne generacije (zamislite par blještavo plavih računala oko 2000. godine, svi postavljeni paralelno, jedan do drugog). Y i mtDNK loze bile su doslovna stabla koja su se u prošlosti konvergirala u jednu točku, a ne u retikulirajuće rešetke. Teorija koalescencije znači kako je svakim korakom unatrag ostajalo sve manje i manje grana za praćenje.
Uspjesi i zamke uniparentalnog doba
Ali, ponekad prilika može biti i prokletstvo. Povijesni populacijski genetičari od prije dvadeset godina, uhvatili su se u zamku gledanja ispod stupa ulične rasvjete. Istraživači su bili duboko zainteresirani za složena pitanja, ali prvi alati koji su im bili na raspolaganju, obiteljska stabla Y i mtDNK, nisu bili dovoljno adekvatni za njihove ambicije.
Kao što smo vidjeli, mtDNK i Y prate samo jednu izravnu liniju genealogije, dok je zbroj demografske povijesti bilo koje populacije gotovo uvijek složena skupina. Ovo ograničenje ponekad se može ilustrirati, čak i putem kontrastnih narativa mtDNK i Y, unutar iste populacije. Npr. plemena Munda u istočno-središnjoj Indiji govore jezikom koji se čini daleki srodnim kambodžanskom. Iako, ako bi gledali samo njihove haplogrupe mtDNK, onda se one savršeno, 100% podudaraju sa svojim indijskim susjedima. Obzirom na ovo, moglo bi se (razumno) odbaciti njihovu jezičnu razliku, i objasniti je samo kao odraz kulturne difuzije. Ali, ako bi pogledali njihove Y-kromosomske loze, otkrili bi kako je 65% njih očito jugoistočnoazijskog podrijetla, a ne indijskog (haplogrupa O, loza koja je vrlo rijetka zapadno od Burme). Dvije linije DNK, majčina i očinska, pričaju vrlo različite priče, priče o dvama potpuno različitim narodima, koji su na kraju isprepleli svoju baštinu u novu populaciju.
Obzirom na miješano stanovništvo Novog svijeta, dobro smo upoznati sa poviješću, pa nakon što potvrdimo kako su mtDNK haplogrupe Argentinaca oko 50% autohtone, nije teško zaključiti kako je Argentina napola autohtona po svom podrijetlu. Svjesni smo kako su generacije europskih migracija, uz iskrivljenu mušku pripadnost, rezultirale uglavnom populacijom europske baštine, čije mtDNK loze ipak pokazuju nedostatak europskih žena među ranim valovima migranata.
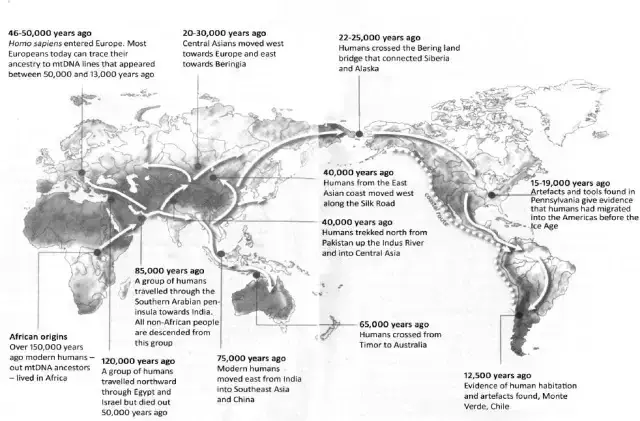
Praćenje ljudske migracije pomoću mtDNK u knjizi "Prava Eva"
Drugi problem je bio u tome što su se djela povijesnog zaključivanja, koja koriste samo suvremene geografske distribucije Y i mtDNK haploskupina, temeljila na prešutnoj pretpostavci kako migracija općenito definira rane faze ljudske ekspanzije. Te, još i gore, kako je nakon toga slijedilo ustaljeno razdoblje relativne stagnacije. Ova pretpostavka je ponekad negdje bila eksplicitno izražena; npr. knjiga Stephena Oppenheimera, "Prava Eva: Putovanje modernog čovjeka izvan Afrike" (2004.), prikazivala je mtDNK filogeniju prekrivenu kartom svijeta (gore), prikazujući pritom sekvencijalnu kolonizaciju planeta tijekom pleistocena, od Afrike do Euroazije, te konačno do Australazije i Novog svijeta. Neka bi se majka naselila u pograničnom području; njene bi kćeri krenule dalje u sljedećoj generaciji - stvarajući mtDNK genealogiju, gdje su genetske, povijesne i geografske udaljenosti bile savršeno usklađene. Razvijajući genealoško stablo, Oppenheimer je predložio čitanje ljudske povijesti u prikazanim obrascima grananja.
Nažalost, čini se kako je osnovna pretpostavka lažna, odnosno tvrdnja kako masovne migracije i promet stanovništva uglavnom prestaju, nakon što ljudi dosegnu određeni teritorij u prapovijesti. To nije problem koji se odnosi isključivo na uniparentalne loze; isto pojednostavljenje mučilo je analize cijelog genoma, i kada su one postale uzlazne.
Kada se ide dalje od jednostavne dinamike brzog širenja i zamjene populacije (npr. migracija iz Afrike, ili zamjena europskih lovaca-sakupljača, odgovaraju tom opisu) - uniparentalni markeri mogu odvesti na krivi put, ili uopće ne sačuvati zapise o skromnijim demografskim promjenama. Nasuprot tome: bogati i gusti podaci, koje nudi analiza cijelog genoma na stotinama tisuća SNP-ova, pružaju bezbrojne filogenetske prozore u povijesti populacije, za razliku od samo dva, te mogu predvidjeti ishode drevne DNK, na načine koja je daleko izvan moći Y i mtDNK. Rad Davida Reicha, naziva "Rekonstrukcija povijesti indijske populacije" (2009.) koristila je moderne uzorke na podacima cijelog genoma, kako bi široko predvidio rezultate u radu iz 2019. godine, "Formiranje ljudskih populacija u Južnoj i Srednjoj Aziji", gdje je koristio stvarne uzorke drevne DNK.
Konačno, tu je problem vremenske dubine i gubitka informacija. Budući se Y i mtDNK loze prenose kroz samo polovicu ljudske populacije, podložne su jačem genetskom driftu i prometu loza. Ispitujući duboku povijest naše vrste, ne možemo se vratiti dalje od najnovijeg zajedničkog pretka, otprilike prije 200000 godina, budući su i Y i mtDNK slijepi izvan ove genetske singularnosti, u onom momentu kada se sve postojeće loze konvergiraju u jednu. To predstavlja problem, jer su se naši neandertalski i denisovski rođaci odvojili od nas prije više od 500000 godina. Sada znamo kako postoji mješavina denisovskih i neandertalskih ljudi, i kod modernih ljudi, ali niti istraživanje Y, niti mtDNK, temeljeno na modernim ljudskim filogenijama nikada nam nije niti dalo naslutiti ovu stvarnost. Ovi pojedinačni markeri jednostavno nemaju široku moć da preusmjere fokus na suptilniju dugoročnu evolucijsku dinamiku naše vrste, zbog njihovih uskih vremenskih horizonta i ograničenja na samo jednu genealogiju.
Dakle, ako su neadekvatni na toliko mnogo načina, zašto se danas spominju ove uniparentalne loze u gotovo svakoj priči o ljudskoj populaciji? Zato jer čak i u toj probnoj fazi, kada je područje pokušavalo učiniti ove ciljane alate istima za sve ljude, nije baš svaki pokušaj bio slijepa ulica. Uspjesi su ilustrirali jedinstvene snage markera, u specifičnim kontekstima, onda kada su evolucijske sile bile toliko sveprisutne i jedna je loza bila dovoljna za hvatanje demografske povijesti; ili kada je mtDNK zbog jednostavnosti ekstrakcije bila najbolji izbor za degradirane uzorke. Jednostavne i elegantne uniparentalne analize su često davale rezultate, koji bi kasnije bili potvrđeni složenijim metodama, onda kada se na internetu pojavilo više računalnih resursa i podataka o sekvenciranju.
Trajna relevantnost mitohondrijske DNK jest naglašena kada je, 2010. godine, pronađena uz niz neolitskih nalazišta u Europi, a šokantni rezultati nagovijestili su ono što će kasnije potvrditi temeljitija genomska drevna DNK: prvi europski poljoprivrednici nisu slični nijednoj drugoj populaciji koja danas živi. Prije 8000 do 5000 godina, kontinentom su gotovo u potpunosti dominirali ljudi uglavnom drevnog anatolijskog podrijetla, netaknuti migracijom iz stepa na istok, a koja će se nakon 3000. godine prije Krista ponovno proširiti kontinentom. Samo su stanovnici Sardinije zadržali svoj neolitski karakter u izolaciji svog otoka. Doseljenici su na njih utjecali tek nakon 1000. godine prije Krista, te i danas nose najveći udio starog poljoprivrednog podrijetla, u cijeloj Europi.

Haplogrupe kao izviđači i tragači
Iz retrospektive, sada možemo vidjeti kako je veliki dio rada na tumačenju distribucije haplogrupa, prije dvadeset godina, mogao biti kao čitanje sudbine iz listića čaja. Uniparentalne loze, same po sebi, jednostavno nisu nudile dovoljno informacija za sastav detaljnih narativa, koji bi bili otporni na buduće obrate i otkrića. U roku od nekoliko godina, moćne nove tehnike bi ih, gotovo u potpunosti, potisnule sa scene.
Ali, do tada su već dramatične "teorije svega", utemeljene samo na Y ili mtDNK filogeniji, bile predložene u popularnim knjigama i dokumentarcima (npr. film "Krv otoka", Bryana Sykesa, iz 2006. godine, zapravo se temeljio samo na malom dijelu krvi otoka). Povijesni genetičari koristili su Y-kromosome i mtDNK kako bi potvrdili da moderni Europljani potječu od pleistocenskih lovaca-sakupljača, što se pokazalo spektakularno pogrešnim, jer lovci-sakupljači nisu ostavili gotovo nikakvo genetsko nasljeđe. Ovo je nesporna činjenica, sada kada možemo usporediti drevnu i modernu DNK. Na Indijskom potkontinentu, prapovjesničari su ignorirali biološke antropološke teorije 20. stoljeća, koje su se temeljile na fizičkom izgledu i koje su tvrdile kako su Južni Azijati mješavina Zapadnih Euroazijaca i autohtonog stanovništva, koji mnogo dulje boravi na tom mjestu. Otpor arheologa i filologa ideji nedavnih masovnih migracija je manje povezana sa vrijednostima teorije, a više je povezano sa idejama njemačkih nacionalista, poput Gustafa Kossinne, koji je postavio temelje za elemente nacističke antropologije. Danas znamo da je stariji antropološki rad uglavnom bio točan u vezi sa Indijom. Zapravo, budimo potpuno pošteni, jer su tada i u genetici postojali glasovi (iako u manjini), koji su ostali otvoreni za migraciju i miješanje. Konačno, iako su genetičari koji se oslanjaju na mtDNK, ispravno naveli kako su Indijci povezani sa populacijama Istočne Azije, potpuno su propustili primjetiti kako je 30-40% njihovih predaka bilo iz paleo-sibirske populacije ("drevni sjeverni Euroazijaci"), sa puno bližim afinitetima prema Europljanima.
I opet, ovo ne znači kako su uniparentalne loze bile beskorisne. Filogenije mtDNK i Y-kromosoma ukorijenjuju podrijetlo modernog čovječanstva unutar Afrike, podupiru dominantno tumačenje fosilnih zapisa, kao i analizu cijelog genoma, tijekom posljednjih dvadeset godina. Kada se integriraju u širu disciplinarnu perspektivu, mogu biti prilično korisne i mogu dosta toga razjasniti, čak pridonijeti odlučujućem tragu, u nekom složenom slučaju. Afiniteti haploskupina Y i mtDNK jasno pokazuju da su Latinoamerikanci miješana populacija, gdje se očinsko i majčino nasljeđe razlikuju, pri čemu prvi potječu s Iberijskog poluotoka, a drugi su autohtoni Južnoamerikanci. Haploskupina Q Y-kromosoma, među autohtonim stanovništvom Sjeverne Amerike i Euroazije je bila nagovještaj dubljih veza između tih populacija, što je kasnije i potvrđeno otkrićem drevnih Sjevernoaurazijaca, ili ANE. Same po sebi, ove uniparentalne loze su tada bile neobjašnjive, ali su od tada postale potpuno razumljivije; npr. dominacija Y-kromosoma kod Q također razotkriva kako je ANE nasljeđe kod Indijanaca snažno iskrivljeno po muškoj liniji.
Pa, čemu onda uniparentalne loze danas najviše doprinose? Pogledajmo nekoliko pokaznih slučajeva.
Jedna od ključnih upotreba jest označavanje mogućih puteva istraživanja. Uzmite u obzir: tijekom 20 godina je detalj distribucije loze R1b predstavljao misterij, tj. zašto regija oko jezera Čad ima toliko visoku učestalost ove uobičajene zapadnoevrozijske haplogrupe? Drevna DNK nam govori kako je visoka učestalost R1b, u zapadnoj Europi, obilježje posljednjih pet tisućljeća migracije. Varijanta R1b, koja je prisutna u Africi, R1b-V88, razlikuje se od svog prekobrojnog europskog rođaka i čini se kao da su se ove dvije grane razišle prije više od 5000 godina. Danas su istraživači, koristeći analizu cijelog genoma, potvrdili tisućljetnu euroazijsku migraciju u središnju Afriku, tako da distribucija ove halpogrupe unutar Afrike više nije takva misterija. Ali, odakle je točno stigao R1b-V88? Drevna DNK nam može odgovoriti na to pitanje; čini se kako potječe iz pretpovijesne Ukrajine. Dubinska i temeljita analiza cijelog genoma male euroazijske frakcije kod stanovnika Čada je mogla donijeti ovaj rezultat uz velike troškove i trud, ali precizne informacije koje je prenijela haplogrupa Y su bile pristupačan i rani prečac do istine.
U najboljem slučaju, nekada imate dovoljno sreće da vam na raspolaganju bude visokokvalitetni cijeli genom. Uz to onda možete nevjerojatno pouzdano rekonstruirati povijest populacije. Stvarnost je često neurednija, pa zahtijeva kreativnu primjenu bilo kakvih alata, koji se trenutno mogu iskoristiti. Prošle godine je objavljen rad o drevnoj DNK, kulture Fatyanovo-Balanovo, koja je cvjetala u zapadnoj Rusiji, i to sjeverno od pontske stepe, prije 4500 godina. Pregledavajući cijele drevne genome, autori su otkrili kako je ova populacija gotovo savršeno odgovarala suvremenim uzorcima kulture Corded Ware sa daljnjeg zapada, kulturi koja je započela u Poljskoj i proširila se prema zapadu, u modernu Njemačku i Skandinaviju. Povijesna lingvistika ispitala je imena rijeka u regiji zapadne Rusije, gdje je Fatyanovo-Balanovo kulturno procvjetalo i pretpostavila kako su ih možda imenovali govornici baltičkog dijalekta (od kojega potječu moderni litavski i latvijski). Ali, u smislu miješanja cijelog genoma, autori nisu mogli doći do nikakvog specifičnog zaključka o afinitetima. Stoga su proučili uniparentalne markere. Mnogi pojedinačni muškarci u grobovima su nosili haplogrupu R1a, koja je danas prevladavajuća među govornicima slavenskih i indoiranskih jezika; uz manji udjel među germanskim i baltičkim skupinama. Među uzorcima, i sa odgovarajućim podacima, šest je imalo Z93 mutaciju R1a. To se gotovo uvijek povezuje sa IndoIrancima, pa su uniparentalni markeri omogućili istraživačima donijeti zaključak kako je kultura i društvo Fatyanovo-Balanovo - bilo jedan od najranijih prethodnika indoiranskih populacija koje su se onda proširile prema istoku, a na kraju i prema jugu, i to tijekom slijedećih 1000 godina. Obzirom na hrpu Y-kromosomskih loza, izoliranih bez pomoći arheološkog, lingvističkog i genomskog konteksta, slučaj bi bio daleko rjeđi. Ali, danas više nije 2000. godina, a pitanjima prapovijesti se može pristupiti iz bezbroj kutova, što daje oštriju stereoskopsku sliku, sve otporniju na izobličenja, kao i čudne vizualne artefakte.
Zatvaranje kruga
Prije jedne generacije, genetika je otišla nezamislivo daleko. Kombinacija znatno moćnijih molekularnih tehnika (poput PCR amplifikacije i ekstrakcije drevne DNK), preciznih dalekosežnih analitičkih okvira (koalescentna teorija), ali i konkretne računalne snage, dala je bogatu informativnu šumu filogenetskih stabala, zasađenih na podacima Y-kromosoma i mtDNK. Koristeći ove nove alate, istraživači su uspostavili razlike između brojnih vrsta i potvrdili afričko podrijetlo cijelog čovječanstva. Zatim su se detaljnije pozabavili ovim zaključcima i utvrdili razlike na razini cijelog kontinenta, u makro haplogrupama, tj. ako je vaša Y haplogrupa bila R, vjerojatno niste bili Istočni Azijat; ako je vaša mtDNK haplogrupa bila U5, vjerojatno ste bili Europljanin. Ali, baš poput Ikara, ubrzo su nadmašili moć svojih tehnika, te su se izgubili u ad hoc i neutemeljenom podudaranju uzoraka. U drugoj polovici 20. stoljeća, genetika se odmaknula od poslovičnog uličnog prikaza Y i mtDNK haplogrupa, i ušla u zasljepljujuća svjetla studija cijelog genoma, gdje su se genetski odnosi na tisućama markera mogli jasno rasvijetliti. I konačno, nakon 2010. godine, drevna DNK počela je davati stvarne genetske podatke iz prošlosti, umjesto samo nagađanja i zaključaka. Bilo je to kao da je sunce izašlo, te sada možemo provoditi ovakva istraživanja pri punom dnevnom svjetlu, a ne u sumračnom talogu.
No, uniparentalni markeri ostaju ključni za razumijevanje povijesti naših predaka i pramajki, natrag do samog početka. Zbog svog spolno pristranog nasljeđivanja, Y i mtDNK opisuju brutalnu stvarnost pretpovijesnog osvajanja. Usporedba Y i mtDNK omogućila je genetičarima zaključiti kako je prije 4000 godina, nekoliko očinskih loza sudjelovalo u masovnoj poliginiji diljem Euroazije, što se podudaralo sa širenjem indoeuropskih jezika. Kombinacija drevne DNK i uniparentalnih markera su također dosada pokazala da su neandertalski Y-kromosomi i mtDNK, zapravo bliži modernim ljudima, nego njihovim denisovskim rođacima. Visokokvalitetni cjeloviti genomi neandertalaca i denisovanaca ne pokazuju jasne rezultate miješanja drevnih modernih ljudi sa neandertalcima, koje nam nude uniparentalni markeri.

Uniparentalni markeri često su vrlo korisni za dobivanje sitnijih detalja, nadograđuju se na procjene ljudske demografske prošlosti, bazirane na razini cijelog genoma. U nekim slučajevima, gdje postoji arheološki promet bez popratnog pomaka na razini cijelog genoma, Y i mtDNK loze još uvijek mogu otkriti promjene. To je slučaj u sjevernoj Europi, gdje je nordijska regija svjedočila višestrukim migracijama sa juga, tijekom 1000 godina. One su se mogle otkriti arheološki, i putem Y-kromosoma, ali ne i putem drugih genetskih analiza. Godine 2800. pr. Kr., migranti koji su potomci kulture kabelske keramike istočne Europe su pristigli u Skandinaviju. Genetski su imali oko 75% stepske baštine, a ostatak je bio od srednjoeuropskih poljoprivrednika, koji su asimilirani neposredno nakon što su nomadi napustili svoju pontsku domovinu. Poput drugih skupina kabelske keramike, svi muškarci među novopridošlima su bili iz Y haplogrupe R1a. Ali, nakon 2500. godine prije Krista, R1a je naglo zamijenjen s R1b, što ukazuje na zamjenu očinske loze, vjerojatno zbog širenja Bell-Beaker sustava sa jugozapada. No, cjelokupni genom izgleda vrlo slično, jer ljudi sa tzv. zvonastim peharima potječu od zapadne populacije, sa vrčastom keramikom. Zatim, nakon 2000. godine prije Krista, učestalost haplogrupe I1, koja ostaje dominantna loza u modernoj Skandinaviji, naglo raste, što ukazuje na još jedan etno-lingvistički pomak, početkom nordijskog brončanog doba. Opet, to je uočljivo samo kod Y-kromosoma, jer se cjelokupno genomsko podrijetlo nije promijenilo. Na svom cjelokupnom genomu, moderni Skandinavci izgledaju otprilike kao potomci populacija sa vrčastom keramikom i ljudi sa zvonastim peharima, ali Y-kromosom priča detaljniju priču. Ovo je klasičan primjer gdje se isplati primijeniti sve naše genetske alate na ova pitanja, jer je naše razumijevanje nepotpuno bez svih gledišta.
Na neki način, tijekom protekle generacije, napravili smo puni krug na uniparentalnim markerima. Kada su prvi put izazvali valove, prije više od jedne generacije, filogenetske analize mtDNK i Y-kromosoma bile su transformativne i kulturno utjecajne, dok su TV dokumentarci najavljivali otkrića i mitohondrijske Eve i Y-kromosomskog Adama. Rješavajući velika pitanja u suradnji sa paleo-antropologijom, uniparentalna populacijska genetika brzo se pokrenula, ispravno utvrdivši cjelokupni oblik nastanka modernog čovječanstva. Problemi su počeli kada su istraživači postali ambiciozniji, rješavajući složena pitanja koja su bila prevelika za ove ciljane i precizne alate. Moći naših nekadašnjih čudaka od djece su bila ograničena, kako onim što sada znamo da su bile pogrešne osnovne pretpostavke o minimalnoj migraciji nakon početnih širenja, tako i inherentnim ograničenjima uske niti naslijeđa koja se proteže kroz tisućljeća.
Iako su se uniparentalni markeri i dalje pokazivali korisnima u procjeni stvari, poput npr. toga koliko je potomaka Džingis-kan mogao imati, do 2000-ih godina se činilo kako je sve više promašaja, a manje pogodaka. Ali, to je bilo tada. Iako uniparentalno podrijetlo odgovara na pitanja tijekom ledenjačke vremenske skale, samo područje paleo-genomike ponekad se može činiti kao da se transformiralo brzinom svjetlosti. Analizi Y i mtDNK odavno su se pridružile dubinske studije cijelih genoma modernih ljudi, kao i ogromno bogatstvo analiza drevne DNK. Dakle, umjesto pretjeranih obećanja ili ambiciozne gradnje na klimavim pretpostavkama, moderna filogenetika Y i mtDNK je pronašla svoju ulogu kao bitan dodatak cjelokupnom alatu sekvenciranja slijedeće generacije, kao i kao dopuna rastućem području paleo-genetike.
Uniparentalni markeri, mtDNK i Y kromosomi, nikada nisu pripadali na glavnu scenu, onda kada su se nakratko pojavili na prijelazu stoljeća. Ali, kako područje paleogenomike nastavlja sazrijevati, oni se nastavljaju sjajno uklapati u rasprostranjenoj sagi naše vrste. Obzirom na to da čak i ljudske stanice vrve mtDNK-om, a Y-kromosome nosi svaki čovjek muškog spola, zapleti oko haplogrupa doslovno su priče o svima nama, o našoj ljudskoj rasi.
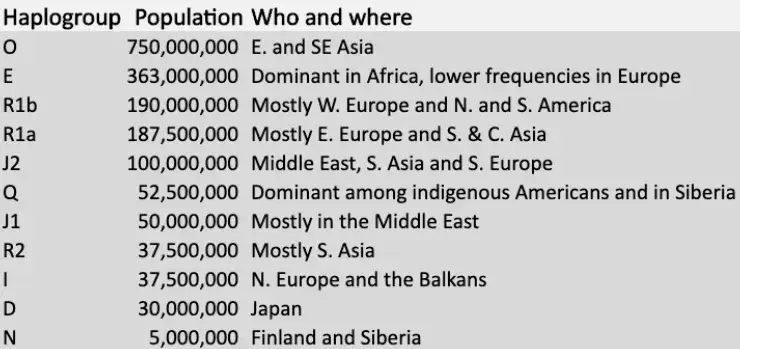
U donjoj tablici pronaći ćete 11 najbrojnijih haplogrupa Y-kromosoma, koje danas postoje među ljudima. Važno je napomenuti da su Y-kromosomi povijesno bili opsesivno proučavani, dijelom i zato što su geografski oštrije odvojeni od mtDNK haplogrupa.
Hvala na čitanju.
Add comment
Comments